
光合成細菌はなぜ放射線を吸収できるのか?
投稿ツリー
-
 放射線の低減に土壌微生物が関係する新たな知見 (go, 2013/9/23 4:30)
放射線の低減に土壌微生物が関係する新たな知見 (go, 2013/9/23 4:30)
-
データ分析表の説明 (go, 2013/9/25 9:14)
-
光合成細菌はなぜ放射線を吸収できるのか? (go, 2013/12/31 8:49)
-

光合成細菌はなぜ放射線を吸収できるのか?
msg# 1.2
go
 投稿数: 125
投稿数: 125
投稿数: 125
「5.考察」では、だいぶ直観的で大胆なことを書いてしまったが、少し検証してみたいと思う。
◆その前に、「微生物で放射能は低減できない」などとネットで発信している人たちがいる。 その根拠は、
① 放射線は、殺菌作用があるので、細菌は生きていけない。
② 光合成細菌が吸収できる光は可視光線近辺だけである。γ放射線は波長が短か過ぎるので吸収できない。
というものらしい
すごく尤もらしい根拠をあげているようだが、間違いも甚だしいですね。
ぜひ一度、私の実証実験のグラフを見ていただきたい、
明らかに「土壌微生物の働きにより放射線が低減している」ことが分かるはずである。
1.「土壌から発するγ放射線は微生物の働きで低減することが明らかになった」と明言した。
グラフを見れば一目瞭然である。以下、
・データ処理は適正に行われていること
・土壌微生物も働きをコントロールしていること
について、整理しながら説明する。
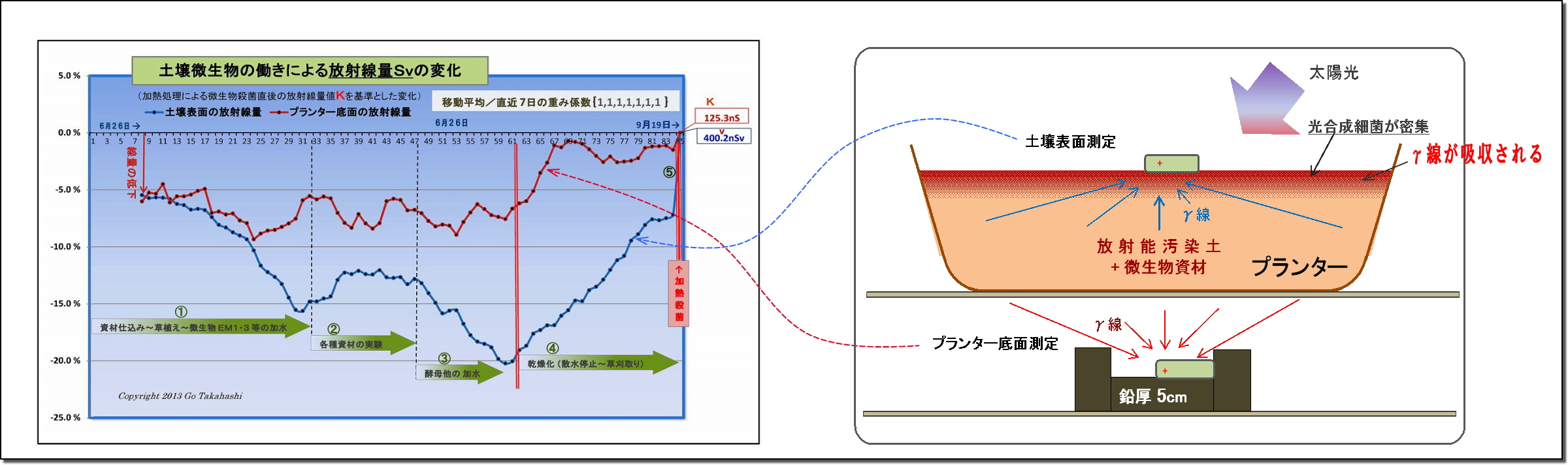
このグラフは、実験プランターの土壌表面の放射線量の変化を示したものである。
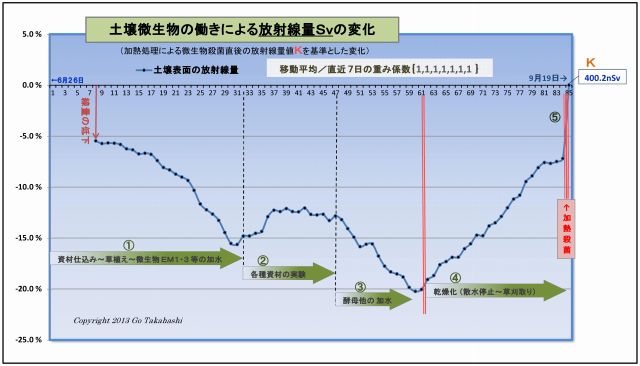
(1) まず最初に、このグラフの元となったデータは適正に処理されている。
・放射線量を連続20点(=200秒)を測定し、それを平均処理し、
・天候等によって変動しがちな空間線量値を岩手県の観測データ値にて補正(減算)し、
・放射能Cs134・Cs137の実験開始からの半減期崩壊値を計算して補正(半減期低下分を加算)し、
・最後に、直前7日間を移動平均して日間のランダム性を平準化した。
このように処理したデータのグラフに見られる放射線量の変化は、
「土壌微生物の働きによるものである」と明言できよう。
(2) 断定できるもう一つの理由「土壌微生物の働きはコントロール」されていることである。
つまり、このデータの変動は偶然に計測さたものではなく、
実験の過程で、微生物の働きを「意図的に操作加減した」結果である、例えば、
・図中①は、実験開始に先立ち、微生物の働きを促進するための微生物と有機資材を汚染土壌と混ぜて、
ヒメイワダレ草も定植したことで光合成細菌をはじめとする土壌微生物叢が豊かになり、
それに伴って、放射線は順調に下がっていること。
・図中②は、放射線の低下が止まり、反転上昇したため、様々な資材(光合成細菌の栄養など)を
試行錯誤的に投入して観察したものであること。
・③は、光合成細菌の働きを良くするために、他の共生微生物を意識的に投入したものである。
例えば「甘酒希釈液」は酵母菌の宝庫であり、この投入で放射線の低下が再び始まり、
③半ばの反転上昇に対しては、「酵母菌増強の発酵液」を散布したこと再び線量は低下した。
この下がった原因は、酵母菌と光合成細菌は共生関係にある、
・光合成細菌がよりいっそう増えるためにはCO2が必要であるが、酵母菌はCO2を排出してくれる。
・一方、酵母菌が増えるためには糖分が必要だが、光合成細菌はブドウ糖を産生する。
・なお、乳酸菌も働くことでphが下がり、腐敗菌が抑えられることで土壌微生物叢が
よりいっそう豊かになる。EM発酵液には乳酸菌などが豊富に含まれている。
・図中④は、資材や水の投入を完全に止め、土壌の乾燥化を促したことによって土壌微生物の働きが弱まり、
その結果として放射線量は上昇(=線量低下が減少)しつづけたこと。
・⑤は、土壌を中華鍋で高熱処理することで水分が皆無となり、
微生物の働きが止ったことで、汚染土壌の本来の放射能によるγ放射線量Kを計測することができ、
このKを基準値として、実験初回からの全データの線量比率を再計算して、上記グラフができあがった。
以上のことから、微生物の働きを意図的にコントロールすることで土壌表面を突き抜けるγ放射線を低減することができた。
(3) 放射線量の変化は、コンプトン散乱によるものではない
実験による放射線量の変化は、放射線等の散乱の影響を受けているのではないか、との指摘はありそうだ。
① 放射線は、電磁波であるとともに光子(粒子性)でもあるので、
強いエネルギーを持つ光子は、物質に衝突すると相互作用を起こすことがある。 例えば、入射した光子が、
・物質の軌道電子をはじき飛ばして消滅する(光電効果)、或いは、
・軌道電子をはじき飛ばし、自らはエネルギーを弱め、方向を変えて散乱する(コンプトン散乱)
そこで下図に、コンプトン散乱についてまとめてみた。
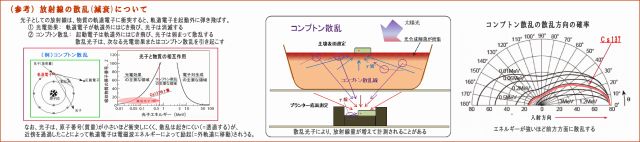
② 入射したγ放射線は、物質との相互作用によって弱められるものであるが、
・しかし、電子番号の低い元素で構成される物質は、ほとんど減衰せずに透過する、
・原子の大きさ(通過断面)に比べて軌道電子の密度が小さいので、光子(粒子)との衝突の確率が少ない。
(原子番号は原子核の陽子の数であり、軌道電子の数でもある)
・微生物を構成する脂質やンパク質は、大半が原子番号の低いNCHOなどで構成されているので、
放射線は軌道電子にほとんど衝突せず、通過することが多い。
③ 今回の実験期間中で変化した物質は、有機資材の投入、土中の草の根の増加だけだが、
これらの増加によって放射線の相互作用が高まり、線量が変化するのだろうか?
・そこで気になるのは、23日目に少量の貝化石を土壌表面にパラパラと撒いたことだが、
その直後の土壌表面線量は、特に目立った変化はなかったので、
コンプトン散乱反射波や光電効果の影響はほとんど無かったものと言えよう。
・他の投入資材は水溶性であるので、プランターの上下全体に行き渡っている。
・草の根は、期間後半は底面に毛根が密集してきけれど、底面線量がそれによって減衰した形跡はみあたらない。
・いずれにしても、光電効果やコンプトン散乱が多少は起こったのだろうけど、
グラフの線量変化は、微生物の働きによって引き起こされたものであることは明らかである。
以上のことから、金属をも透過するほどのエネルギーの強いγ線であっても、
・土壌微生物は放射線を吸収している
・微生物の量と活性度合いで吸収量は増加する
という結論に行きつく。
なお参考までに、フクイチ原発事故以来、福島県はもとより北関東や宮城県などにおいて、
EMを定期的に散布することで「放射線量が下がる」ことが、フォーラムやネットなどで多く報告されているが、
私の実証実験の結果からみても、納得できるものであることを付記しておきたい。
2.「γ放射線の低減は光合成細菌の働きによるものと思われる」
これは、次の根拠によるものである。
・放射線量は、土壌表面の方がプランター底面よりも低下(=吸収)が大きいこと。
・光合成細菌は、太陽光の当たる土壌表面に密集すること。
・光子エネルギーを利用吸収できる微生物は光合成細菌だけであること。
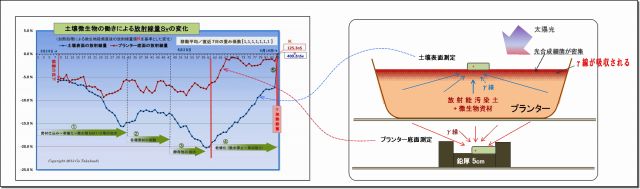
(1) γ放射線の物質の透過力は非常に強い
・Cs137のγ放射線は、可視光線と同じ光子ではあるが、可視光線より遥かに強い(30万倍)エネルギーを持っている。
・今回の実験では、土壌表面では、最大20%ほど放射線量が低下しているが、
この低下の量は、厚さ2mmの鉛板で遮蔽した時の減衰量に匹敵する。
・期間を通して、土壌表面と測定器を遮るものは特にない。
(厳密に言えば、草はあるけど、測定器の検出部は草の茎のない処に置いているし、
試しに草の茎や葉を少し寄せ集めてその上から計ってみたが測定データは変わらなかった)
(2) 土壌表面のγ線量の低下は光合成細菌の仕業と思われる
① 光合成細菌の密度は土壌表面の方が遥かに高い
・光合成細菌は、太陽光からエネルギーを受けやすい表面近くに密集(=増殖)する。
このことにより、徐々にではあるが、土中(底面方向)より遥かに密度が高くなっていった。
・実験開始直後のしばらくは、プランター底面も土壌表面も低下率は同じであったが、
微生物叢が活発になりはじめると、次第に土壌表面の方が低下率が大きくなったが、その理由については、
光合成細菌が土壌表面に多く集まってきて、放射線を底面よりも多く吸収していると考える。
(3) 鉛板2mmに相当する放射線の減衰は、微生物タンパクなる単なる物質による減衰であるとは到底考え難い。
・物質による減衰ではなく、「生命体として積極的に吸収している現象」と考えたい、
「吸収」とい表現よりも、「エネルギーを積極的に取り込んでいる」と言った方が正確ではないだろうか。
・プランター内の生体は土壌微生物だけであり、光子エネルギーを利用できるのは生体は光合成細菌しかない。
消去法ではあるが、放射線の光子エネルギーを吸収しているのは光合成細菌である、と断言したい。
光合成細菌が、その密度を高め、活動が活性化するほど、放射線の吸収量は増加する、ということである。
しかし、この「光合成細菌の放射線吸収説」を裏付けるには、難儀するねぇ・・・、
ネットのどこを探しても「光合成細菌はγ放射線を吸収する」という情報は見当たらないのだ
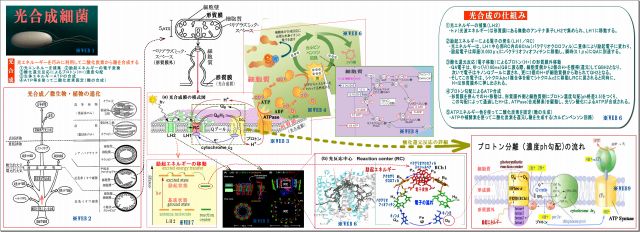
3. 光合成細菌による光合成のメカニズム
そこで、光合成細菌がγ線を吸収できるという根拠を探るため、光合成のメカニズムをまとめてみた。
結論を先に言うなら、
・光合成細菌の基質(=エサ)は、光(光子エネルギー)と二酸化炭素である、これを原料として糖を合成する。
・γ放射線は、可視光線よりも遥かに強い光子エネルギーもっている。
*WEB1 *WEB2 *WEB3 *WEB4 *WEB5 *WEB6 *WEB7 *WEB8 *WEB9
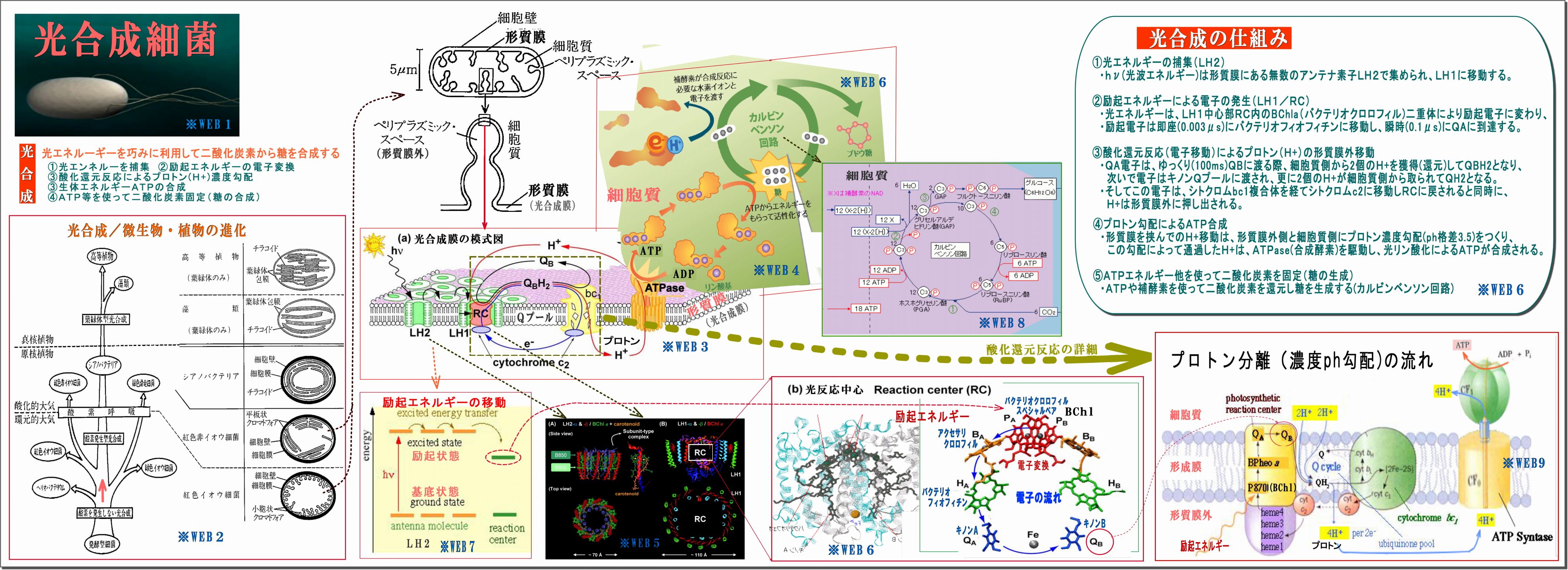
(1) 光合成細菌の細胞膜(形質膜:光合成膜とも言う)には光合成機能が詰まっている
・細胞膜が細胞内に向って小胞上のくびれ(クロマトフォア)がたくさんある。
この図では紅色イオウ細菌のものだが、紅色非イオウ細菌の場合は、クロマトフォアが
細胞質内に更にツリー状に広がっている。
・そのクロマトフォアには、たくさんの「光合成機能セット」が詰まっている、図はその一セットを拡大したもので、
特に光合成膜の面積大半を占めるのは、光エネルギー取り込むアンテナ素子LH2である。
(2) 光合成とは、光エネルギーを巧みに利用して二酸化炭素から糖を合成すること。 その流れは、
① 光エネルギーを捕集
・光で励起された光エネルギーをできるだけ広く集めるためのアンテナ素子がLH2であり、
・その励起エネルギーはLH1の反応中心RC(Reaction Center)に向って伝達される。
・励起エネルギーの伝搬方法は、励起とは軌道電子がより高い軌道に移動することであるが、
その電子は、瞬時に元の軌道に戻る際に他の原子の軌道電子を励起するという。
・この励起エネルギーはLH2→LH1→RCに瞬時に伝搬するらしい。
② 励起エネルギーの電子変換
・励起エネルギーはRCの中核BChl(バクテリオクロロフィル)にて電子に変換され、
バクテリオフィオフィチン→キノンQAへと瞬時に伝達される
③ 酸化還元反応によるプロトン(H+)濃度勾配
・QA電子はQBおよびキノンQプールに移動する際、細胞質側からプロトンH+を取ってQH2となり、
・電子は、後でシトクロム複合体を経てシトクロムc2からRCのヘムに戻されるが、
その前に、H+は形質膜外に押し出され、形質膜の内外にプロトン濃度(ph)勾配ができる。
④ 生体エネルギーATPの合成
・このプロトン濃度(ph)勾配によって、H+はATPase(合成酵素)を通過することで駆動され、
細胞質側で光リン酸化によってATPが合成される。
⑤ ATPを使って二酸化炭素固定(糖の合成)
・細胞質内では、ATPや補酵素を使って二酸化炭素を還元し、糖を合成する(カルビンベンソン回路)
3.光合成細菌が吸収できるのは可視光線だけなのか?
そんなことはない。
・いまや光合成細菌の吸収スペクトルは、研究され公表されているが、
この吸収スペクトルは、可視光線を照射して特定した「光学的スペクトル」のことであり、
光子エネルギー全般について特定されたものではない。
・光合成細菌は、光子エネルギーを吸収するメカニズムが備わっており、
γ放射線やX線などの光子エネルギーをも利用することができる。
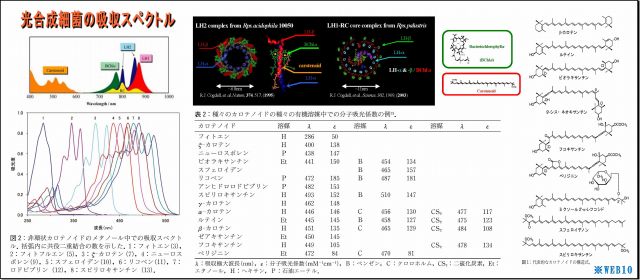
*WEB10
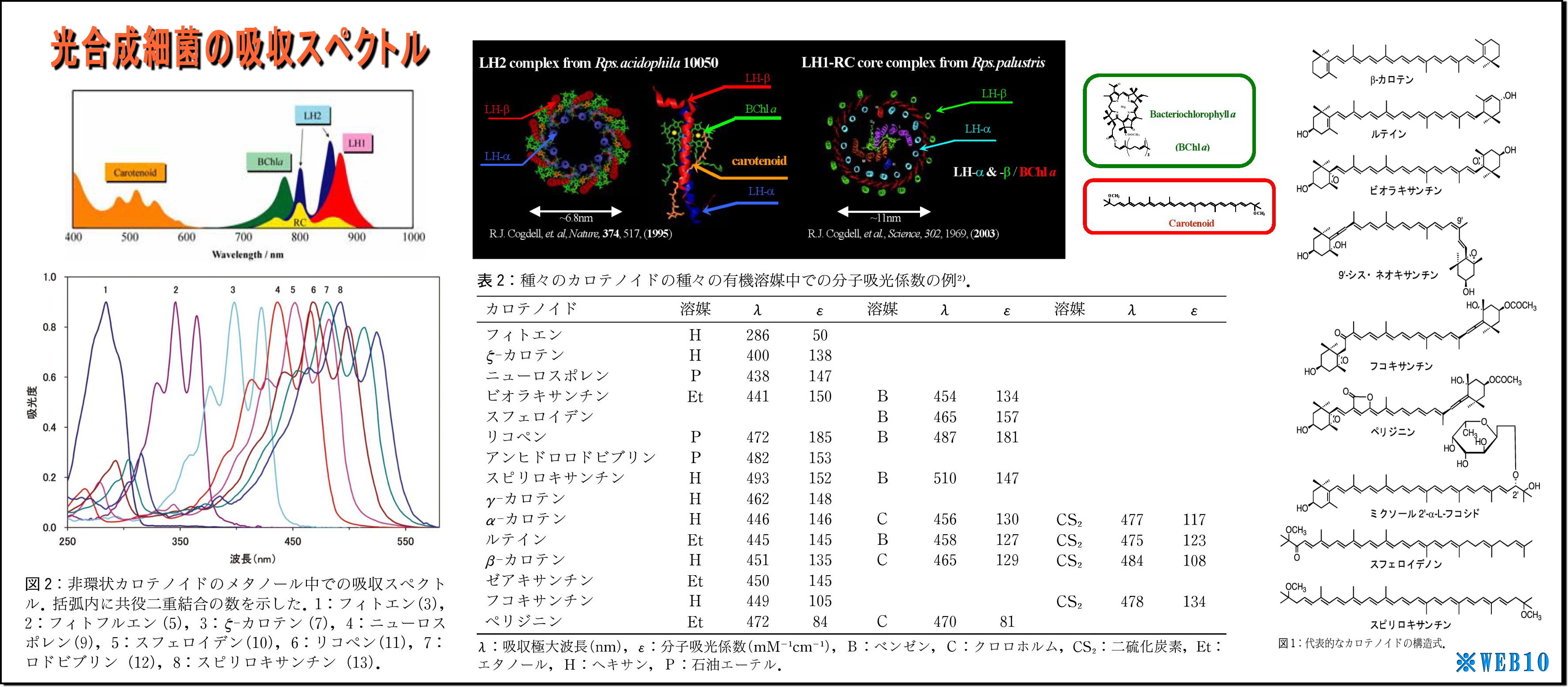
(1) LH2の立体構造を見ると、ヘリカル(螺旋)構造が目についてくる
・これは光も電磁波なので、光を効率よく集めるためなのだろうか、実に見事な形状である、
・また、このLH2の直径は70Å(オングストローム、1Å=0.1nm)=7nm程度であり、ヘリカル径はもっと小さいことから、
可視光線波長(390~750nm)向けアンテナ形状にしては小さいような気もする。
・そもそも原子の大きさは数nmなので、波長と共振できる形状だけから言及するなら、
分子レベルの大きさと構造(すなわち原子配列・形状・光入射向き)や色素複合体などとの関係が重要なのだろう。
・図の「吸収スペクトル」をみると、
・分子構造が複雑で大きいほど赤外線などの長波長に対応しているようであり、
・集光エネルギー終着点であるBChl(バクテリオクロロフィル)もスペクトルを持つが、
これはもっぱら光励起エネルギーの電子変換が主な役割らしい。
・またこれに絡み付くように繋がっているカロチノイドは短波長に対応するという、
・カロチノイドの基幹である共益二重結合の数Nが少ないほど短波長を吸収するようで、
N=3のフィトンでも280nm付近(UV-B中紫外線)止まりのようだ。
以上のことから、
吸収(エネルギー利用)できる光の範囲は、近紫外線~近赤外線(波長300~1,000nm)であるとされており、、
波長0.00187nmであるCs137のγ放射線は、吸収スペクトルから大きく外れていることになる。
このことをもって、「光合成細菌は放射線を吸収できない」「利用されない」と断言する人もいるようだ、
が、果たして、そう断言できるのか?
(2) しかし、私の実験では「γ放射線は光合成細菌によって吸収されている」という実証データがある。
① 公表されたスペクトル論に異を唱えるつもりは毛頭ない。
しかし、もっと短い波長の光子は吸収できない、とは書かれていない。
(3) 研究として公表されている「吸収スペクトル」の特定方法を見ると、
① タンパク質を溶媒に抽出し、近赤外線~可視光線~近紫外線の光を当てて調べたものである。
・つまり、単なる「光学的特性」の結果でしかない、ということだ。
・光は電磁波でもあるので、吸収スペクトルとは、本来電子回路系をも含めた周波数特性のことである。
電子回路でいうLCR(Lコイル、Cコンデンサ、R抵抗)のインピーダンスと周波数との共振性であり、
アンテナとは密接に関係するけど、決してアンテナ素子の物質・形状だけで決まるものではない。
・しかも、電子回路としてみた場合、一連の光化学プロセスは実に厄介な構造であり、いまだ解明されない処も
随所にあることから、周波数スペクトルを特定するには、実際に電子を流して検証する必要がある。
しかし、それは一切なされていない。
②更には、実際にX線や放射線を当てて吸収スペクトルを調べた訳ではもちろんない。
・地球に降り注ぐ太陽エネルギーの大半は可視光線とその前後だから対象外なのだろう。
(参考までに)いまの太陽光線中の割合は、およそ、
遠紫外線1%以下、近紫外線8%程度、可視光線40%、近赤外線20%、中赤外線25%、遠赤外線6% らしい。
・それとも、放射線の強い透過力では、吸収の変化は計測不能に近いからなのだろうか・・・。
・なお、LHCのヘリカル構造の直径は数Åであり、何やらX線帯近くまで対応できているような気がするが。
③そもそも、アンテナ素子LHC(Lh2、Lh1)は、光子エネルギーを受けて電子励起するものである。
・光子エネルギーは何も可視光に限らない、遠紫外線~X線~放射線のエネルギーは、
可視光の数百倍~数万倍~従十万倍もの強い光子エネルギーを持っているものなので、
放射線がLHCを通過する(軌道電子と衝突する確率は低い)だけで容易に軌道電子は励起されよう。
・つまり、アンテナ素子の元素配列や形状(光学的スペクトル)とは無関係に励起される。
・太古の地球では地表に降り注ぐ太陽エネルギーは陽子線・中性子線、X線や放射線が多かった事からして、
その時代に出現した光合成細菌は、生いたち初期からエネルギーの強い放射線やX線を利用していたと思われる。
LHCのヘリカル構造や分子の元素構造や配列形状は、放射線やXが少なくなる時期と合わせて進化したのだろう、
実に巧みに設計されたものと思われる。
④ 放射線の照射で細菌は死滅するということへの反論
・エネルギーMeV以上の強い放射線を局所集中的に照射するなら、死滅すると思われる。
・LHC構造のカロチノイドには、強い照射に対する防御機能(活性酸素を打ち消す役割)もあるらしい
・私の実験では、光合成細菌や酵母などの共生菌が死滅または増殖が阻害されているとは考えられない、
むしろ元気なくらいに感じられる。
・チェルノブイリ事故周辺やフクシマ現場周辺の写真を見る限り、植物が赤茶けて枯れた様子は全く見られない。
・光合成の備わった生物は、放射能に強く、光合成も盛んになるもの、と思われる。
⑤ なお、X線や放射線の吸収性(スペクトル)を確かめたいならば、
・細菌が死んだ状態では計れない、単に透過するだけだから、生きてシステムが働いている状態で測定しなければならない。
・なお、例え細菌が生きていたとしても、負荷が必要である、負荷とは出力負荷=消費のことだ、
つまり、合成されたATPを消費してあげないと、飽和して励起エネルギーの流れはすぐに停滞してしまう、
・電気実験だって、いくら電圧をかけても電力が消費されなければ電流は流れない。
・ATPが消費され、合成された糖などが共生菌によって消費され、更に植物草に消費される、など、
一連の微生物叢や植物との共生が出来上がって、はじめて放射線の吸収が確認できるものと考えたい。
この観点から、私の実験では、詳細なスペクトルまでは調べることはできないが、
間違いなく「放射線を吸収している」ことがを実証しているのである。
4. 光合成細菌の放射線吸収の謎にせまる
光合成細菌は、太古の地球が太陽風(陽子線・放射線・紫外線に晒されていた環境で出現し、進化してきた微生物なので、
放射線に強いというよりも「放射線を利用できるメカニズム」が備わっているものと推測する。
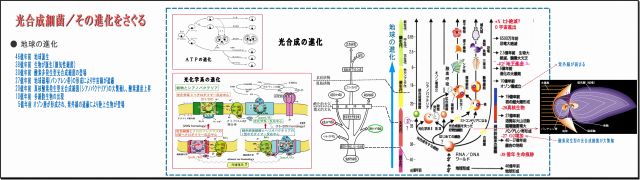
① 光合成細菌に光エネルギー捕集と電子変換の見事なメカニズム
・実に見事に設計されているものと感嘆する。
(イ) 電磁波エネルギーと光子エネルギーの両方を利用できるメカニズムとなっている
(ロ) 電磁波エネルギーは、広く配置したアンテナ素子LHCで捕集する機能、
可能な限り広範囲な可視光線帯のエネルギーを吸収しやすいために
クロロフィルや多様な分子構造をもつカロチノイドの構造と配置によるスペクトルの形成
(ハ) 可視光線より遥かに波長の短い光子(遠紫外線・X線・γ線)のエネルギーは、
光学的スペクトルとは無関係に、直接LHCを励起する。
(ニ) 可視光線近辺の光捕集による励起エネルギーも、より短波長の光子による励起エネルギーも、
バクテリオクロロフィルPにて同一視して電子変換されてる
(ホ) 電子のキノンと結びつき、ATP合成のためのプロトン勾配を作る
(ヘ) 生体エネルギーATPを利用して糖を合成する
・実に見事としか言いようがない。
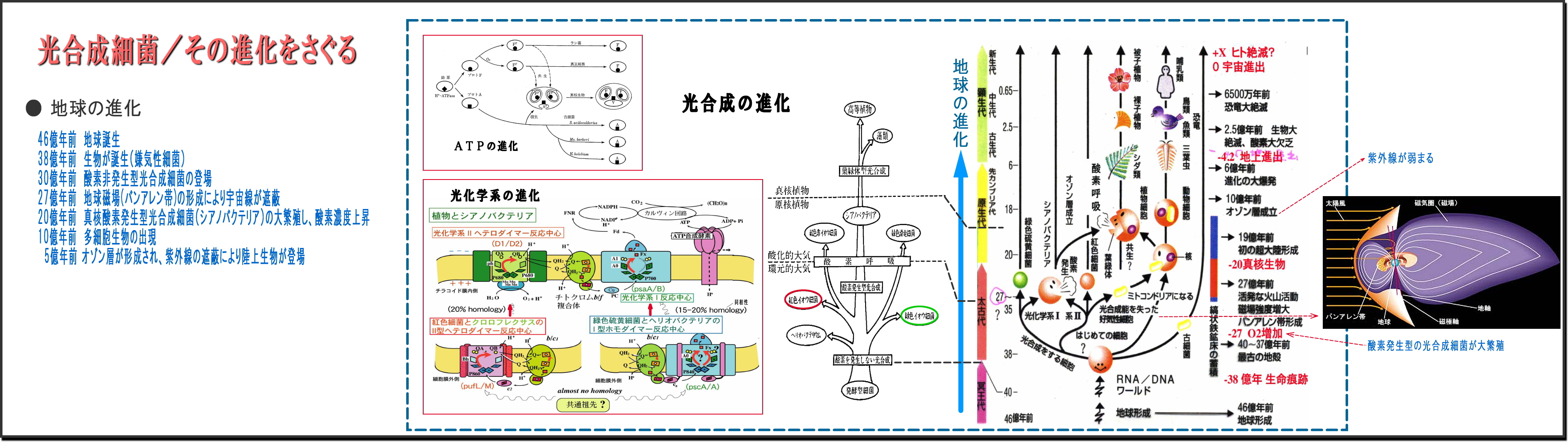
② 地球の進化は生物の進化と共にある
・地球という惑星が誕生したのは46億年前で、最初の生命体誕生は、今なお謎らしい。
38億年前の岩石には、すでに生命の痕跡が見つかっている。
・最初の生物は微生物だが、他から栄養をもらって増殖する従属栄養微生物が多いなか、
自ら栄養を作り出せる独立栄養微生物は光合成細菌だけである、
・この光合成細菌の誕生(微生物の進化)によって、地球は飛躍的に生命力が増してきた。
・光合成細菌の最初は、酸素を発生しない光合成細菌であった。
有毒ガスである硫化水素の多い地球だから、H2Sを還元して光化学する紅色(や緑色)硫黄細菌であり、
次第にH2Oを還元し酸素を排出する非硫黄細菌へと進化する。
・この光合成細菌の光合成機能を受け継いだ原核植物シアノバクテリア(藍藻)へと進化し、
ついには植物の葉のなかの葉緑体へと発展していく。
・この進化のようすはクロマトフォアの密集度合いの推移にみることができる。
・なお余談だが、光合成の中間産物である生体エネルギーATP機能は、動物細胞のミトコンドリア
として細胞内に組込まれていく。
③ 太古の地球には酸素はほとんど無く、やがて活発なマグマが落ち着くころになると、
・地殻が安定し、自転による地磁気が明確になってバンアレン帯が形成される27億年前ころになると、
陽子線や放射線などが含まれる有害な地球への太陽風は次第に弱まり、
やがて酸素の層が厚みを帯びてきて、10億年前頃にはオゾン層ができると紫外線も弱まり、
太陽エネルギーは可視光線がメインとなってきた。
④ 光合成細菌は、このバンアレン帯が出来上がる以前に登場し進化を続けてきた
・バンアレン帯以前の地球は、陽子線や放射線など生体に有害エネルギーが降り注ぐ状態である、
こんな環境でも、光合成細菌のタンパク質などは破壊されないで進化・増殖できたのには、
放射線をも基質(増殖に必要なエサ)として利用してきたと想像する。
・可視光線の数十万倍ものエネルギーのある放射線を、生命の増殖に使わない手はないし、
過酷な環境に順応し、逆に過酷な環境を好むように光合成細菌は進化してきたものと素直に考えたい。
⑤ 光合成細菌の進化について
・上記①に述べたように、光合成の見事なまでの光化学メカニズムは、いきなり完成したわけではない。
・複雑な光学的スペクトルが形成できたのは、進化のずっと後のことで、放射線よりも可視光線の割合が
多くなってきたことに適応しながらの進化ではなかろうか。
・つまり、地球太古の時代は、可視光線よりも遥かにエネルギー高いX線やγ線が降り注いでいたので、
簡単に励起エネルギーを得ることができるシンプルな光化学系メカニズムが最初にできあがった、と考えたい。
そして、バンアレン帯以降あたりから可視光線をも吸収しやすいような光学スペクトル分子構造へと進化してきた。
以上、私がいろいろ調べてみたことを、私の推測も交えて、備忘録的に取りまとめてはみた。
だいぶ謎の核心に近づいてきたのではないかと思っているが、如何なものだろうか
◆その前に、「微生物で放射能は低減できない」などとネットで発信している人たちがいる。 その根拠は、
① 放射線は、殺菌作用があるので、細菌は生きていけない。
② 光合成細菌が吸収できる光は可視光線近辺だけである。γ放射線は波長が短か過ぎるので吸収できない。
というものらしい
すごく尤もらしい根拠をあげているようだが、間違いも甚だしいですね。
ぜひ一度、私の実証実験のグラフを見ていただきたい、
明らかに「土壌微生物の働きにより放射線が低減している」ことが分かるはずである。
1.「土壌から発するγ放射線は微生物の働きで低減することが明らかになった」と明言した。
グラフを見れば一目瞭然である。以下、
・データ処理は適正に行われていること
・土壌微生物も働きをコントロールしていること
について、整理しながら説明する。
このグラフは、実験プランターの土壌表面の放射線量の変化を示したものである。
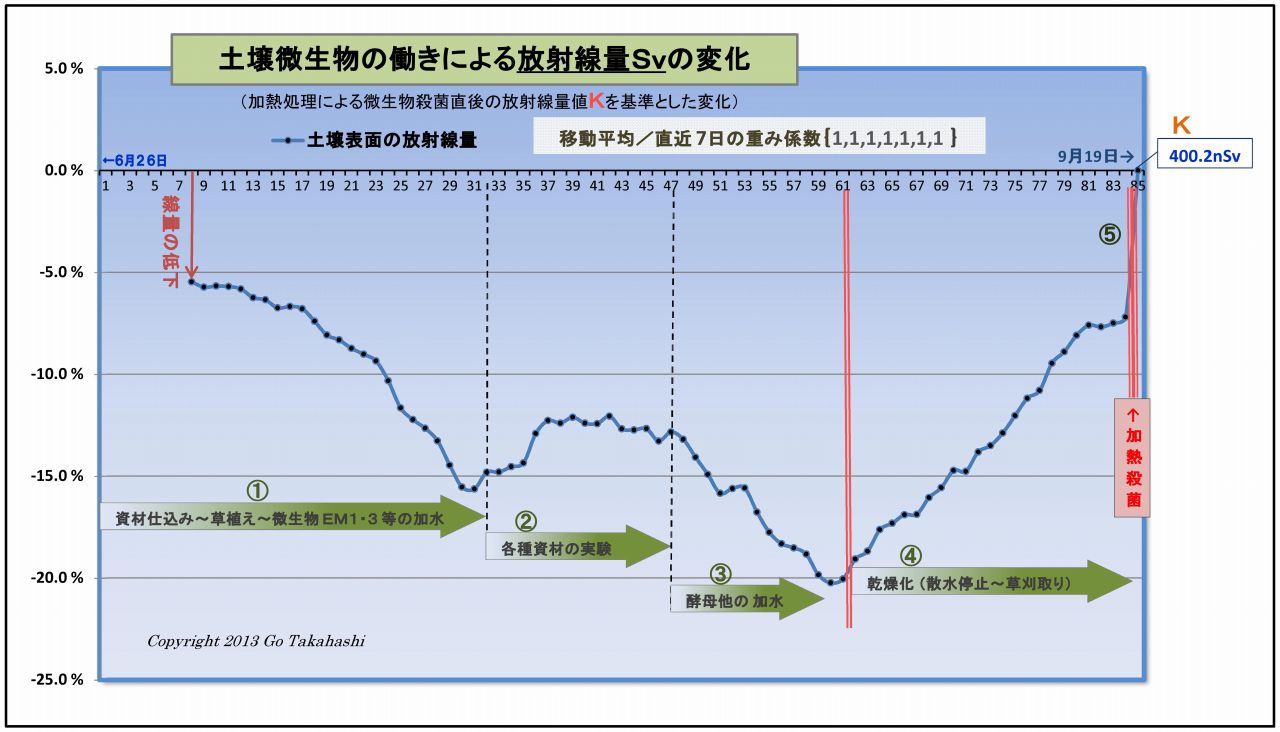
(1) まず最初に、このグラフの元となったデータは適正に処理されている。
・放射線量を連続20点(=200秒)を測定し、それを平均処理し、
・天候等によって変動しがちな空間線量値を岩手県の観測データ値にて補正(減算)し、
・放射能Cs134・Cs137の実験開始からの半減期崩壊値を計算して補正(半減期低下分を加算)し、
・最後に、直前7日間を移動平均して日間のランダム性を平準化した。
このように処理したデータのグラフに見られる放射線量の変化は、
「土壌微生物の働きによるものである」と明言できよう。
(2) 断定できるもう一つの理由「土壌微生物の働きはコントロール」されていることである。
つまり、このデータの変動は偶然に計測さたものではなく、
実験の過程で、微生物の働きを「意図的に操作加減した」結果である、例えば、
・図中①は、実験開始に先立ち、微生物の働きを促進するための微生物と有機資材を汚染土壌と混ぜて、
ヒメイワダレ草も定植したことで光合成細菌をはじめとする土壌微生物叢が豊かになり、
それに伴って、放射線は順調に下がっていること。
・図中②は、放射線の低下が止まり、反転上昇したため、様々な資材(光合成細菌の栄養など)を
試行錯誤的に投入して観察したものであること。
・③は、光合成細菌の働きを良くするために、他の共生微生物を意識的に投入したものである。
例えば「甘酒希釈液」は酵母菌の宝庫であり、この投入で放射線の低下が再び始まり、
③半ばの反転上昇に対しては、「酵母菌増強の発酵液」を散布したこと再び線量は低下した。
この下がった原因は、酵母菌と光合成細菌は共生関係にある、
・光合成細菌がよりいっそう増えるためにはCO2が必要であるが、酵母菌はCO2を排出してくれる。
・一方、酵母菌が増えるためには糖分が必要だが、光合成細菌はブドウ糖を産生する。
・なお、乳酸菌も働くことでphが下がり、腐敗菌が抑えられることで土壌微生物叢が
よりいっそう豊かになる。EM発酵液には乳酸菌などが豊富に含まれている。
・図中④は、資材や水の投入を完全に止め、土壌の乾燥化を促したことによって土壌微生物の働きが弱まり、
その結果として放射線量は上昇(=線量低下が減少)しつづけたこと。
・⑤は、土壌を中華鍋で高熱処理することで水分が皆無となり、
微生物の働きが止ったことで、汚染土壌の本来の放射能によるγ放射線量Kを計測することができ、
このKを基準値として、実験初回からの全データの線量比率を再計算して、上記グラフができあがった。
以上のことから、微生物の働きを意図的にコントロールすることで土壌表面を突き抜けるγ放射線を低減することができた。
(3) 放射線量の変化は、コンプトン散乱によるものではない
実験による放射線量の変化は、放射線等の散乱の影響を受けているのではないか、との指摘はありそうだ。
① 放射線は、電磁波であるとともに光子(粒子性)でもあるので、
強いエネルギーを持つ光子は、物質に衝突すると相互作用を起こすことがある。 例えば、入射した光子が、
・物質の軌道電子をはじき飛ばして消滅する(光電効果)、或いは、
・軌道電子をはじき飛ばし、自らはエネルギーを弱め、方向を変えて散乱する(コンプトン散乱)
そこで下図に、コンプトン散乱についてまとめてみた。
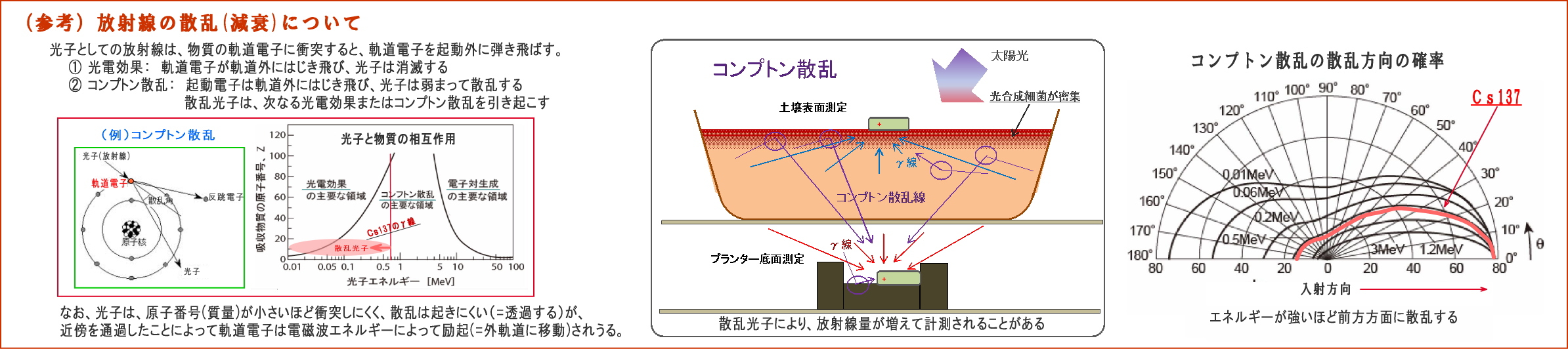
② 入射したγ放射線は、物質との相互作用によって弱められるものであるが、
・しかし、電子番号の低い元素で構成される物質は、ほとんど減衰せずに透過する、
・原子の大きさ(通過断面)に比べて軌道電子の密度が小さいので、光子(粒子)との衝突の確率が少ない。
(原子番号は原子核の陽子の数であり、軌道電子の数でもある)
・微生物を構成する脂質やンパク質は、大半が原子番号の低いNCHOなどで構成されているので、
放射線は軌道電子にほとんど衝突せず、通過することが多い。
③ 今回の実験期間中で変化した物質は、有機資材の投入、土中の草の根の増加だけだが、
これらの増加によって放射線の相互作用が高まり、線量が変化するのだろうか?
・そこで気になるのは、23日目に少量の貝化石を土壌表面にパラパラと撒いたことだが、
その直後の土壌表面線量は、特に目立った変化はなかったので、
コンプトン散乱反射波や光電効果の影響はほとんど無かったものと言えよう。
・他の投入資材は水溶性であるので、プランターの上下全体に行き渡っている。
・草の根は、期間後半は底面に毛根が密集してきけれど、底面線量がそれによって減衰した形跡はみあたらない。
・いずれにしても、光電効果やコンプトン散乱が多少は起こったのだろうけど、
グラフの線量変化は、微生物の働きによって引き起こされたものであることは明らかである。
以上のことから、金属をも透過するほどのエネルギーの強いγ線であっても、
・土壌微生物は放射線を吸収している
・微生物の量と活性度合いで吸収量は増加する
という結論に行きつく。
なお参考までに、フクイチ原発事故以来、福島県はもとより北関東や宮城県などにおいて、
EMを定期的に散布することで「放射線量が下がる」ことが、フォーラムやネットなどで多く報告されているが、
私の実証実験の結果からみても、納得できるものであることを付記しておきたい。
2.「γ放射線の低減は光合成細菌の働きによるものと思われる」
これは、次の根拠によるものである。
・放射線量は、土壌表面の方がプランター底面よりも低下(=吸収)が大きいこと。
・光合成細菌は、太陽光の当たる土壌表面に密集すること。
・光子エネルギーを利用吸収できる微生物は光合成細菌だけであること。
(1) γ放射線の物質の透過力は非常に強い
・Cs137のγ放射線は、可視光線と同じ光子ではあるが、可視光線より遥かに強い(30万倍)エネルギーを持っている。
・今回の実験では、土壌表面では、最大20%ほど放射線量が低下しているが、
この低下の量は、厚さ2mmの鉛板で遮蔽した時の減衰量に匹敵する。
・期間を通して、土壌表面と測定器を遮るものは特にない。
(厳密に言えば、草はあるけど、測定器の検出部は草の茎のない処に置いているし、
試しに草の茎や葉を少し寄せ集めてその上から計ってみたが測定データは変わらなかった)
(2) 土壌表面のγ線量の低下は光合成細菌の仕業と思われる
① 光合成細菌の密度は土壌表面の方が遥かに高い
・光合成細菌は、太陽光からエネルギーを受けやすい表面近くに密集(=増殖)する。
このことにより、徐々にではあるが、土中(底面方向)より遥かに密度が高くなっていった。
・実験開始直後のしばらくは、プランター底面も土壌表面も低下率は同じであったが、
微生物叢が活発になりはじめると、次第に土壌表面の方が低下率が大きくなったが、その理由については、
光合成細菌が土壌表面に多く集まってきて、放射線を底面よりも多く吸収していると考える。
(3) 鉛板2mmに相当する放射線の減衰は、微生物タンパクなる単なる物質による減衰であるとは到底考え難い。
・物質による減衰ではなく、「生命体として積極的に吸収している現象」と考えたい、
「吸収」とい表現よりも、「エネルギーを積極的に取り込んでいる」と言った方が正確ではないだろうか。
・プランター内の生体は土壌微生物だけであり、光子エネルギーを利用できるのは生体は光合成細菌しかない。
消去法ではあるが、放射線の光子エネルギーを吸収しているのは光合成細菌である、と断言したい。
光合成細菌が、その密度を高め、活動が活性化するほど、放射線の吸収量は増加する、ということである。
しかし、この「光合成細菌の放射線吸収説」を裏付けるには、難儀するねぇ・・・、
ネットのどこを探しても「光合成細菌はγ放射線を吸収する」という情報は見当たらないのだ
3. 光合成細菌による光合成のメカニズム
そこで、光合成細菌がγ線を吸収できるという根拠を探るため、光合成のメカニズムをまとめてみた。
結論を先に言うなら、
・光合成細菌の基質(=エサ)は、光(光子エネルギー)と二酸化炭素である、これを原料として糖を合成する。
・γ放射線は、可視光線よりも遥かに強い光子エネルギーもっている。
*WEB1 *WEB2 *WEB3 *WEB4 *WEB5 *WEB6 *WEB7 *WEB8 *WEB9
(1) 光合成細菌の細胞膜(形質膜:光合成膜とも言う)には光合成機能が詰まっている
・細胞膜が細胞内に向って小胞上のくびれ(クロマトフォア)がたくさんある。
この図では紅色イオウ細菌のものだが、紅色非イオウ細菌の場合は、クロマトフォアが
細胞質内に更にツリー状に広がっている。
・そのクロマトフォアには、たくさんの「光合成機能セット」が詰まっている、図はその一セットを拡大したもので、
特に光合成膜の面積大半を占めるのは、光エネルギー取り込むアンテナ素子LH2である。
(2) 光合成とは、光エネルギーを巧みに利用して二酸化炭素から糖を合成すること。 その流れは、
① 光エネルギーを捕集
・光で励起された光エネルギーをできるだけ広く集めるためのアンテナ素子がLH2であり、
・その励起エネルギーはLH1の反応中心RC(Reaction Center)に向って伝達される。
・励起エネルギーの伝搬方法は、励起とは軌道電子がより高い軌道に移動することであるが、
その電子は、瞬時に元の軌道に戻る際に他の原子の軌道電子を励起するという。
・この励起エネルギーはLH2→LH1→RCに瞬時に伝搬するらしい。
② 励起エネルギーの電子変換
・励起エネルギーはRCの中核BChl(バクテリオクロロフィル)にて電子に変換され、
バクテリオフィオフィチン→キノンQAへと瞬時に伝達される
③ 酸化還元反応によるプロトン(H+)濃度勾配
・QA電子はQBおよびキノンQプールに移動する際、細胞質側からプロトンH+を取ってQH2となり、
・電子は、後でシトクロム複合体を経てシトクロムc2からRCのヘムに戻されるが、
その前に、H+は形質膜外に押し出され、形質膜の内外にプロトン濃度(ph)勾配ができる。
④ 生体エネルギーATPの合成
・このプロトン濃度(ph)勾配によって、H+はATPase(合成酵素)を通過することで駆動され、
細胞質側で光リン酸化によってATPが合成される。
⑤ ATPを使って二酸化炭素固定(糖の合成)
・細胞質内では、ATPや補酵素を使って二酸化炭素を還元し、糖を合成する(カルビンベンソン回路)
3.光合成細菌が吸収できるのは可視光線だけなのか?
そんなことはない。
・いまや光合成細菌の吸収スペクトルは、研究され公表されているが、
この吸収スペクトルは、可視光線を照射して特定した「光学的スペクトル」のことであり、
光子エネルギー全般について特定されたものではない。
・光合成細菌は、光子エネルギーを吸収するメカニズムが備わっており、
γ放射線やX線などの光子エネルギーをも利用することができる。
*WEB10
(1) LH2の立体構造を見ると、ヘリカル(螺旋)構造が目についてくる
・これは光も電磁波なので、光を効率よく集めるためなのだろうか、実に見事な形状である、
・また、このLH2の直径は70Å(オングストローム、1Å=0.1nm)=7nm程度であり、ヘリカル径はもっと小さいことから、
可視光線波長(390~750nm)向けアンテナ形状にしては小さいような気もする。
・そもそも原子の大きさは数nmなので、波長と共振できる形状だけから言及するなら、
分子レベルの大きさと構造(すなわち原子配列・形状・光入射向き)や色素複合体などとの関係が重要なのだろう。
・図の「吸収スペクトル」をみると、
・分子構造が複雑で大きいほど赤外線などの長波長に対応しているようであり、
・集光エネルギー終着点であるBChl(バクテリオクロロフィル)もスペクトルを持つが、
これはもっぱら光励起エネルギーの電子変換が主な役割らしい。
・またこれに絡み付くように繋がっているカロチノイドは短波長に対応するという、
・カロチノイドの基幹である共益二重結合の数Nが少ないほど短波長を吸収するようで、
N=3のフィトンでも280nm付近(UV-B中紫外線)止まりのようだ。
以上のことから、
吸収(エネルギー利用)できる光の範囲は、近紫外線~近赤外線(波長300~1,000nm)であるとされており、、
波長0.00187nmであるCs137のγ放射線は、吸収スペクトルから大きく外れていることになる。
このことをもって、「光合成細菌は放射線を吸収できない」「利用されない」と断言する人もいるようだ、
が、果たして、そう断言できるのか?
(2) しかし、私の実験では「γ放射線は光合成細菌によって吸収されている」という実証データがある。
① 公表されたスペクトル論に異を唱えるつもりは毛頭ない。
しかし、もっと短い波長の光子は吸収できない、とは書かれていない。
(3) 研究として公表されている「吸収スペクトル」の特定方法を見ると、
① タンパク質を溶媒に抽出し、近赤外線~可視光線~近紫外線の光を当てて調べたものである。
・つまり、単なる「光学的特性」の結果でしかない、ということだ。
・光は電磁波でもあるので、吸収スペクトルとは、本来電子回路系をも含めた周波数特性のことである。
電子回路でいうLCR(Lコイル、Cコンデンサ、R抵抗)のインピーダンスと周波数との共振性であり、
アンテナとは密接に関係するけど、決してアンテナ素子の物質・形状だけで決まるものではない。
・しかも、電子回路としてみた場合、一連の光化学プロセスは実に厄介な構造であり、いまだ解明されない処も
随所にあることから、周波数スペクトルを特定するには、実際に電子を流して検証する必要がある。
しかし、それは一切なされていない。
②更には、実際にX線や放射線を当てて吸収スペクトルを調べた訳ではもちろんない。
・地球に降り注ぐ太陽エネルギーの大半は可視光線とその前後だから対象外なのだろう。
(参考までに)いまの太陽光線中の割合は、およそ、
遠紫外線1%以下、近紫外線8%程度、可視光線40%、近赤外線20%、中赤外線25%、遠赤外線6% らしい。
・それとも、放射線の強い透過力では、吸収の変化は計測不能に近いからなのだろうか・・・。
・なお、LHCのヘリカル構造の直径は数Åであり、何やらX線帯近くまで対応できているような気がするが。
③そもそも、アンテナ素子LHC(Lh2、Lh1)は、光子エネルギーを受けて電子励起するものである。
・光子エネルギーは何も可視光に限らない、遠紫外線~X線~放射線のエネルギーは、
可視光の数百倍~数万倍~従十万倍もの強い光子エネルギーを持っているものなので、
放射線がLHCを通過する(軌道電子と衝突する確率は低い)だけで容易に軌道電子は励起されよう。
・つまり、アンテナ素子の元素配列や形状(光学的スペクトル)とは無関係に励起される。
・太古の地球では地表に降り注ぐ太陽エネルギーは陽子線・中性子線、X線や放射線が多かった事からして、
その時代に出現した光合成細菌は、生いたち初期からエネルギーの強い放射線やX線を利用していたと思われる。
LHCのヘリカル構造や分子の元素構造や配列形状は、放射線やXが少なくなる時期と合わせて進化したのだろう、
実に巧みに設計されたものと思われる。
④ 放射線の照射で細菌は死滅するということへの反論
・エネルギーMeV以上の強い放射線を局所集中的に照射するなら、死滅すると思われる。
・LHC構造のカロチノイドには、強い照射に対する防御機能(活性酸素を打ち消す役割)もあるらしい
・私の実験では、光合成細菌や酵母などの共生菌が死滅または増殖が阻害されているとは考えられない、
むしろ元気なくらいに感じられる。
・チェルノブイリ事故周辺やフクシマ現場周辺の写真を見る限り、植物が赤茶けて枯れた様子は全く見られない。
・光合成の備わった生物は、放射能に強く、光合成も盛んになるもの、と思われる。
⑤ なお、X線や放射線の吸収性(スペクトル)を確かめたいならば、
・細菌が死んだ状態では計れない、単に透過するだけだから、生きてシステムが働いている状態で測定しなければならない。
・なお、例え細菌が生きていたとしても、負荷が必要である、負荷とは出力負荷=消費のことだ、
つまり、合成されたATPを消費してあげないと、飽和して励起エネルギーの流れはすぐに停滞してしまう、
・電気実験だって、いくら電圧をかけても電力が消費されなければ電流は流れない。
・ATPが消費され、合成された糖などが共生菌によって消費され、更に植物草に消費される、など、
一連の微生物叢や植物との共生が出来上がって、はじめて放射線の吸収が確認できるものと考えたい。
この観点から、私の実験では、詳細なスペクトルまでは調べることはできないが、
間違いなく「放射線を吸収している」ことがを実証しているのである。
4. 光合成細菌の放射線吸収の謎にせまる
光合成細菌は、太古の地球が太陽風(陽子線・放射線・紫外線に晒されていた環境で出現し、進化してきた微生物なので、
放射線に強いというよりも「放射線を利用できるメカニズム」が備わっているものと推測する。
① 光合成細菌に光エネルギー捕集と電子変換の見事なメカニズム
・実に見事に設計されているものと感嘆する。
(イ) 電磁波エネルギーと光子エネルギーの両方を利用できるメカニズムとなっている
(ロ) 電磁波エネルギーは、広く配置したアンテナ素子LHCで捕集する機能、
可能な限り広範囲な可視光線帯のエネルギーを吸収しやすいために
クロロフィルや多様な分子構造をもつカロチノイドの構造と配置によるスペクトルの形成
(ハ) 可視光線より遥かに波長の短い光子(遠紫外線・X線・γ線)のエネルギーは、
光学的スペクトルとは無関係に、直接LHCを励起する。
(ニ) 可視光線近辺の光捕集による励起エネルギーも、より短波長の光子による励起エネルギーも、
バクテリオクロロフィルPにて同一視して電子変換されてる
(ホ) 電子のキノンと結びつき、ATP合成のためのプロトン勾配を作る
(ヘ) 生体エネルギーATPを利用して糖を合成する
・実に見事としか言いようがない。
② 地球の進化は生物の進化と共にある
・地球という惑星が誕生したのは46億年前で、最初の生命体誕生は、今なお謎らしい。
38億年前の岩石には、すでに生命の痕跡が見つかっている。
・最初の生物は微生物だが、他から栄養をもらって増殖する従属栄養微生物が多いなか、
自ら栄養を作り出せる独立栄養微生物は光合成細菌だけである、
・この光合成細菌の誕生(微生物の進化)によって、地球は飛躍的に生命力が増してきた。
・光合成細菌の最初は、酸素を発生しない光合成細菌であった。
有毒ガスである硫化水素の多い地球だから、H2Sを還元して光化学する紅色(や緑色)硫黄細菌であり、
次第にH2Oを還元し酸素を排出する非硫黄細菌へと進化する。
・この光合成細菌の光合成機能を受け継いだ原核植物シアノバクテリア(藍藻)へと進化し、
ついには植物の葉のなかの葉緑体へと発展していく。
・この進化のようすはクロマトフォアの密集度合いの推移にみることができる。
・なお余談だが、光合成の中間産物である生体エネルギーATP機能は、動物細胞のミトコンドリア
として細胞内に組込まれていく。
③ 太古の地球には酸素はほとんど無く、やがて活発なマグマが落ち着くころになると、
・地殻が安定し、自転による地磁気が明確になってバンアレン帯が形成される27億年前ころになると、
陽子線や放射線などが含まれる有害な地球への太陽風は次第に弱まり、
やがて酸素の層が厚みを帯びてきて、10億年前頃にはオゾン層ができると紫外線も弱まり、
太陽エネルギーは可視光線がメインとなってきた。
④ 光合成細菌は、このバンアレン帯が出来上がる以前に登場し進化を続けてきた
・バンアレン帯以前の地球は、陽子線や放射線など生体に有害エネルギーが降り注ぐ状態である、
こんな環境でも、光合成細菌のタンパク質などは破壊されないで進化・増殖できたのには、
放射線をも基質(増殖に必要なエサ)として利用してきたと想像する。
・可視光線の数十万倍ものエネルギーのある放射線を、生命の増殖に使わない手はないし、
過酷な環境に順応し、逆に過酷な環境を好むように光合成細菌は進化してきたものと素直に考えたい。
⑤ 光合成細菌の進化について
・上記①に述べたように、光合成の見事なまでの光化学メカニズムは、いきなり完成したわけではない。
・複雑な光学的スペクトルが形成できたのは、進化のずっと後のことで、放射線よりも可視光線の割合が
多くなってきたことに適応しながらの進化ではなかろうか。
・つまり、地球太古の時代は、可視光線よりも遥かにエネルギー高いX線やγ線が降り注いでいたので、
簡単に励起エネルギーを得ることができるシンプルな光化学系メカニズムが最初にできあがった、と考えたい。
そして、バンアレン帯以降あたりから可視光線をも吸収しやすいような光学スペクトル分子構造へと進化してきた。
以上、私がいろいろ調べてみたことを、私の推測も交えて、備忘録的に取りまとめてはみた。
だいぶ謎の核心に近づいてきたのではないかと思っているが、如何なものだろうか